細胞壁
細胞壁(英語:)是植物細胞的外層,在細胞膜的外面,細胞壁之厚薄常因組織、功能不同而異。它可以是坚韧的,有弹性,和有时坚硬的。它提供细胞既有结构支撐和保护,例如將動物細胞放入純水,會進水量比出水量多而造成爆破;但植物細胞因為有細胞壁,頂多只會膨脹,不至爆破。細胞壁同时也作为一种过滤机制。植物、真菌(菌物)、藻類和原核生物都具有細胞壁,而支原体属細胞不具有細胞壁。

细胞壁的组成随着不同物种而变化,并可能取决于细胞的类型和发展阶段。陆生植物的初生细胞壁()的组成是多糖类的纤维素,半纤维素和果胶。在细菌中,细胞壁的组成是肽聚糖。古菌细胞壁有各种组分物组成,并可能由糖蛋白的S层,假肽聚糖或多糖组成的。真菌具有N-乙酰葡萄糖胺的聚合物幾丁質组成的细胞壁,和藻类通常具有糖蛋白和多糖组成的细胞壁。与众不同的是,矽藻具有一个由生物硅组成的细胞壁[1]。其他辅助分子往往也锚定到细胞壁中,例如木质素和几丁质。
成分與功能
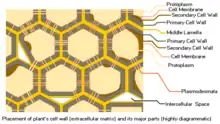
細胞壁是在細胞分裂、生長和分化過程中形成的,主要成分是纤维素和果胶,可用于支撐和維持植物細胞的形狀。細胞壁分為三層,初生細胞壁()和次生細胞壁()。細胞與細胞之間有胞間層()分隔。所有植物細胞都有初生壁,其外面就是胞間層。次生壁在初生壁的裡面,是在細胞停止生長後分泌形成的,可以增加細胞壁的厚度和強度,不易受到病原物多醣降解酶的直接攻擊,但不是所有的細胞都具有次生壁。
次生壁又分內(S3)、中(S2)、外(S1)三層(在內層裡面,有時還可出現一層),外層和內層都很薄,只有中層最厚,佔次生壁厚度的70~90%,主要成分也是半纤维素、纤维素和木質素(lignin),極少含果膠,久之會開始進行不同程度的木質化,木聚醣逐漸分佈於整個次生壁中,而木葡聚醣則局限分佈於初生壁和胞間層,另外,角質(cutin)和木栓質(suberin)通常會埋入次生壁中。次生壁越厚,壁內的細胞腔就越小,等到細胞完全成熟後細胞腔呈現橢圓形。
植物的細胞壁
植物細胞的壁必須具有足夠的抗拉強度,以承受幾倍大氣壓的內部滲透壓,這是由細胞內部溶液和外部溶液之間的溶質濃度差異引起的[2]。 植物細胞壁的厚度在0.1到幾μm之間變化[3]。
分層
在植物細胞壁中可以發現多達三個層[4]:
- 初生細胞壁,通常是在細胞生長時形成的薄的,柔性的和可延伸的層。
- 次生細胞壁,在細胞完全生長後在初生細胞壁內形成的厚層。 在所有細胞類型中均已發現。 一些細胞,例如木質部中的傳導細胞,具有含有木質素的第二壁,使細胞壁增強並防水。
- 中膠層(Middle layer),一層富含果膠。 該最外層形成相鄰植物細胞之間的界面並將它們粘合在一起。
成分
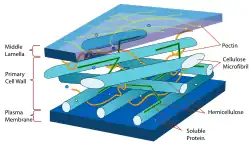
在初生(生長)植物細胞壁中,主要的碳水化合物是纖維素,半纖維素和果膠。 纖維素微纖維通過半纖維素系鏈連接以形成纖維素 - 半纖維素網絡,其嵌入果膠基質中。 在初生細胞壁中最常見的半纖維素是木葡聚醣[5]。在草的細胞壁中,木葡聚醣和果膠的豐度減少,部分被另一種半纖維素的葡醣醛酸阿拉伯木聚醣取代。 原代細胞壁特徵性地通過稱為酸生長的機制延伸(生長),由擴張蛋白介導,由酸性條件激活的細胞外蛋白質,其修飾果膠和纖維素之間的氫鍵[6]。這起到增加細胞壁延伸性的作用。 植物表皮的初生細胞壁的外部通常用角質和蠟浸漬,形成稱為植物角質層的滲透性屏障。
次生細胞壁(Secondary cell walls)含有多種其他化合物,可改變其機械性能和滲透性能。 構成木材(主要是次生細胞壁)的主要聚合物包括:
此外,在大多數植物細胞壁中發現結構蛋白(1-5%); 它們被分類為富含羥脯氨酸的糖蛋白(HRGP),阿拉伯半乳聚醣蛋白(AGP),富含甘氨酸的蛋白質(GRP)和富含脯氨酸的蛋白質(PRPs)。每一類的糖蛋白是被一個特徵性的高度重複的蛋白質序列來定義。 大多數是被糖基化的,含有羥脯氨酸(Hyp)並在細胞壁中交聯。 這些蛋白質通常集中在特化細胞和細胞角落中。表皮的細胞壁可含有角質。 根部內皮層中的凯氏带(Casparian strip)和植物樹皮的木栓細胞含有木栓質。 角質素和木栓質都是聚酯,其作用是水的運動的滲透屏障[7]。碳水化合物,次級化合物和蛋白質的相對組成在植物之間以及細胞類型和年齡之間變化。 植物細胞壁還含有許多酶,例如水解酶,酯酶,過氧化物酶和轉糖基酶(transglycosylases),其作用是切割,修剪和交叉鏈接細胞壁聚合物。
真菌的細胞壁

有幾組生物被稱為“真菌”。 這些組中的一些(卵菌綱和Myxogastria)已被轉移出真菌域,部分原因是細胞壁組成的基本生化差異。 大多數真正的真菌都有細胞壁,主要由幾丁質和其他多醣組成[9]。 真正的真菌在細胞壁中沒有纖維素[10]。
其他真核細胞壁
藻類

像植物一樣,藻類有細胞壁[12]。 藻類細胞壁含有多醣(例如纖維素(一种葡聚醣))或多種糖蛋白(团藻目)或兩者都有。 在藻類細胞壁中包含額外的多醣被用作藻類分類的特徵。
- 甘露聚醣:它們在許多海洋綠藻的細胞壁中形成微纖維,包括來自Codium,绒枝藻属和傘藻屬的那些屬,以及一些紅藻的細胞壁,例如紫菜属(Porphyra)和红毛菜属(Bengia)。
- 木聚醣:
- 海藻酸:它是褐藻細胞壁中常見的多醣。
- 磺化的多醣:它們存在於大多數藻類的細胞壁中; 紅藻中常見的包括洋菜,卡拉胶,紫菜属,红藻胶(furcelleran),和海萝胶(funoran)。
在藻類細胞壁中可能積累的其他化合物包括孢粉素和鈣離子。
原核生物的細胞壁
細菌細胞壁
細胞膜外側是細菌細胞壁。 細菌細胞壁由肽聚醣製成,其由多醣鏈製成,所述多醣鏈由含有D-氨基酸的不尋常肽交聯[16]。細菌細胞壁不同於分別由纖維素和幾丁質製成的植物和真菌的細胞壁[17]。細菌的細胞壁也不同於不含肽聚醣的古菌細胞壁。儘管L型细菌可以在缺乏細胞壁的實驗室中產生,但細胞壁對許多細菌的存活至關重要[18]。抗生素青黴素能夠通過阻止肽聚醣的交聯來殺死細菌,這會導致細胞壁變弱和溶解[17]。 溶菌酶也可以破壞細菌細胞壁。
從廣義上講,細菌中有兩種不同類型的細胞壁,革蘭氏陰性和革蘭氏陽性。這些名稱源於細胞對革蘭氏染色的反應,這是一種長期用於細菌種類分類的試驗[19]。
革蘭氏陰性菌的細胞壁的結構和組成複雜,肽聚糖只有一薄層,在兩層細胞膜之間的膜間腔中,而革蘭氏陽性菌細胞壁厚得多,且只含有很多層肽聚糖。
各個生物界細胞壁的差異
参考文献
- Rutledge, Ryan D.; Wright, David W. . Lukehart, C.M.; Scott, R.A. (编). . EIC Books. Wiley. 2013 [2016-03-14]. ISBN 978-1-118-62522-4.
- . [2018-07-14]. (原始内容存档于2021-04-03).
- Campbell, Neil A.; Reece, Jane B.; Urry, Lisa A.; Cain, Michael L.; Wasserman, Steven A.; Minorsky, Peter V.; Jackson, Robert B. 8th. 2008: 118. ISBN 978-0-8053-6844-4.
- Buchanan; Gruissem, Jones. 1st. American society of plant physiology. 2000. ISBN 0-943088-39-9.
- Fry, Stephen C. . Journal of Experimental Botany. 1989, 40 (1): 1–11. doi:10.1093/jxb/40.1.1.
- Braidwood, Luke; Breuer, Christian; Sugimoto, Keiko. . New Phytologist. 2013-08-29, 201 (2): 388–402 [2018-07-14]. ISSN 0028-646X. doi:10.1111/nph.12473. (原始内容存档于2020-07-23) (英语).
- Laurence Moire; Alain Schmutz; Antony Buchala; Bin Yan; Ruth E. the; Ulrich Ryser. . Plant Physiol. 1999, 119 (3): 1137–1146 [2018-07-17]. PMC 32096
 . PMID 10069853. doi:10.1104/pp.119.3.1137. (原始内容存档于2010-11-11).
. PMID 10069853. doi:10.1104/pp.119.3.1137. (原始内容存档于2010-11-11). - Jarvis, Michael C. . Plant Physiology. 2013-12-01, 163 (4): 1485–1486 [2018-07-19]. PMC 3850196 . PMID 24296786. doi:10.1104/pp.113.231092. (原始内容存档于2021-04-11).
- Hudler, George W. (1998). Magical Mushrooms, Mischievous Molds. Princeton, NJ: Princeton University Press, 7. ISBN 0-691-02873-7.
- Webster, John & Weber, Roland W.S. (2007) "Introduction to Fungi" New York, NY: Cambridge University Press, 6."
- Webster, John & Weber, Roland W.S. (2007) "Introduction to Fungi" New York, NY: Cambridge University Press, 5-7."
- Sendbusch, Peter V. (2003-07-31). "Cell Walls of Algae 的存檔,存档日期November 28, 2005,.". Botany Online. Retrieved on 2007-10-29.
- Sengbusch, Peter V. (2003-07-31). "Interactions between Plants and Fungi: the Evolution of their Parasitic and Symbiotic Relations 的存檔,存档日期December 8, 2006,.". biologie.uni-hamburg.de. Retrieved on 2007-10-29.
- Alexopoulos, C. J., C. W. Mims, & M. Blackwell (1996). Introductory Mycology 4. New York: John Wiley & Sons, 687-688. ISBN 0-471-52229-5.
- Raper, Kenneth B. (1984). The Dictyostelids. Princeton, NJ: Princeton University Press, 99-100. ISBN 0-691-08345-2.
- van Heijenoort J. . Glycobiology. 2001, 11 (3): 25R – 36R [2018-07-17]. PMID 11320055. doi:10.1093/glycob/11.3.25R. (原始内容存档于2009-07-01).
- Koch A. . Clin Microbiol Rev. 2003, 16 (4): 673–87 [2018-07-17]. PMC 207114 . PMID 14557293. doi:10.1128/CMR.16.4.673-687.2003. (原始内容存档于2008-09-28).
- Joseleau-Petit D, Liébart JC, Ayala JA, D'Ari R. . J. Bacteriol. September 2007, 189 (18): 6512–20 [2018-07-17]. PMC 2045188 . PMID 17586646. doi:10.1128/JB.00273-07. (原始内容存档于2019-10-18).
- Gram, HC. . Fortschr. Med. 1884, 2: 185–189.
- White, David. (1995) The Physiology and Biochemistry of Prokaryotes, pages 6, 12-21. (Oxford: Oxford University Press). ISBN 0-19-508439-X.