原生質絲
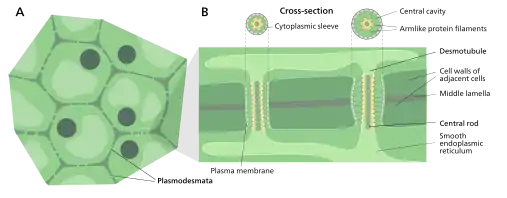
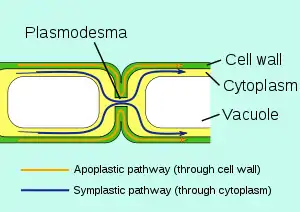
原生質絲(英語:)為植物細胞和部分藻類細胞壁間貫穿細胞壁的特有孔道,可以讓相鄰細胞的細胞質相互流通。[2][3][4]有微小孔道,為細胞間物質運輸與信息傳遞的重要通道,通道中有一連接兩細胞內質網的連絲微管,細胞質可經由原生質絲交流及運輸,此過程稱為共質體運輸。
目前已知擁有原生質絲的包含植物界的所有物種,以及藻類中輪藻綱、輪藻目、褐藻綱和鞘毛藻目。[5]
植物細胞擁有動物細胞所沒有的細胞壁,鄰近的細胞藉由薄板相互分隔。雖然細胞壁對於部分蛋白質及溶質具有穿透性,但原生質絲對於共質體內的胞內運輸更可以控制原生質流的流向以及穿透的物質。
原生質絲有分兩種:初生原生質絲(primary plasmodesmata)及次生原生質絲(secondary plasmodesmata)。初生原生質絲在細胞分裂期間出現,而次生原生質絲是真正溝通成熟植物細胞的通道。[6]
動物也有類似的構造,允許動物細胞間的物質交流,包含間隙連接(gap junctions)[7]和膜奈米管(membrane nanotubes)[8][9]
組成
原生質絲是當植物細胞分裂時,部份的高基氏體釋出的囊泡附於中膠層上。分裂時,中膠層上會產生一些小孔,可以溝通原生質,即為初生原生質絲,而在植物細胞成熟後,細胞壁逐漸加厚,這些小孔便會成為一條一條的管狀構造,即為次級原生質絲。[10]
構造
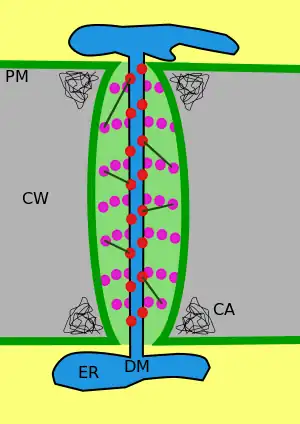
原生質絲壁上的細胞膜
一個典型的植物細胞通常有103至105條原生質絲。[11][12]胞質套筒的寬度約50-60 nm。
原生質絲壁可分為三層主層:細胞膜、胞質套筒(cytoplasmic sleeve)以及連絲微管(desmotubule)[11],它們可以穿過約90 nm的細胞壁[12]。
運輸
原生質絲可以讓蛋白質(包括轉錄因子)、小干擾RNA、mRNA和病毒的基因組在細胞內流通。其中一個病毒的運動蛋白例子是MP-30。MP-30可結合病毒的基因組,並且將病毒的基因組經由原生質絲傳入未受感染的細胞。[14]Flowering Locus T protein也可經由原生質絲從葉部移動至頂芽分生組織以促使開花。[21]
可經由原生質絲傳送的粒子大小並不一定,植物可藉由活躍的調整機制來控制通過顆粒的大小。[6]而相對的某些顆粒也可經由改變自己的大小來穿越原生質絲,如MP-30的大小可在700道耳吞到9400道耳吞之間,藉以協助某些物質傳送到整株植物。[22]
學者已提出數種原生質絲的模型,認為原生質絲通透性的調節,是藉由與蛋白質或是部分未經折疊的伴隨蛋白的交互作用而調整。[23]
註釋
- Maule, Andrew. . Current Opinion in Plant Biology. December 2008, 11 (6): 680–686. PMID 18824402. doi:10.1016/j.pbi.2008.08.002.
- Oparka, K. J. (2005) Plasmodesmata. Blackwell Pub Professional. ISBN 1-4051-2554-3 ISBN 978-1-4051-2554-3
- . [2016-02-15]. (原始内容存档于2016-02-22).
- (PDF). [2015-11-12]. (原始内容存档 (PDF)于2016-07-29).
- Graham, LE; Cook, ME; Busse, JS (2000), Proceedings of the National Academy of Sciences 97, 4535-4540.
- http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1692983 The shoot apical meristem: the dynamics of a stable structure. Jan Traas and Teva Vernoux : Philos Trans R Soc Lond B Biol Sci. 2002 June 29; 357(1422): 737–747. (page 744)
- Bruce Alberts. 4th. New York: Garland Science. 2002. ISBN 0-8153-3218-1.
- Gallagher KL, Benfey PN. . Genes Dev. January 2005, 19 (2): 189–95 [2013-08-12]. PMID 15655108. doi:10.1101/gad.1271005. (原始内容存档于2019-09-12).
- Gray JC, Sullivan JA, Hibberd JM, Hansen MR. . Plant Biology. 2001, 3: 223–33. doi:10.1055/s-2001-15204.
- Lucas W., Ding, B. and Van der Schoot, C. (1993) Tansley Review No.58 "Plasmodesmata and the supracellular Nature of Plants" New Phytologist, Vol. 125, No. 3, pp. 435-476, Stable URL:http://www.jstor.org/stable/2558257 (页面存档备份,存于)
- AW Robards (1975) Plasmodesmata. Annual Review of Plant Physiology 26, 13-29
- Lodish, Berk, Zipursky, Matsudaira, Baltimore, Darnell. . 4. 2000: 998. ISBN 0-7167-3706-X.
- AW Robards (1976) Plasmodesmata in higher plants. In: Intercellular communications in plants: studies on plasmodesmata. Edited by BES Gunning and AW Robards Springer-Verlag Berlin pps 15-57.
- Plasmodesmata and the control of symplastic transport (页面存档备份,存于) A. G. ROBERTS & K. J. OPARKA
- RL Overall, J Wolfe, BES Gunning (1982) Intercellular communication in Azolla roots: I. Ultrastructure of plasmodesmata. Protoplasma 111: 134-150
- LC Cantrill, RL Overall and PB Goodwin (1999) Cell-to-cell communication via plant endomembranes. Cell Biology International 23: 653–661
- JE Radford and RG White (1998) Localization of a myosin‐like protein to plasmodesmata. Plant Journal 14: 743-750
- LM Blackman and RL Overall (1998) Immunolocalisation of the cytoskeleton to plasmodesmata of Chara corallina. Plant Journal 14: 733-741
- S Reichelt, AE Knight, TP Hodge, F Baluska, J Samaj, D Volkmann and J Kendrick-Jones (1999) Characterization of the unconventional myosin VIII in plant cells and its localization at the post-cytokinetic cell wall. Plant Journal 19: 555–569
- RG White, K Badelt, RL Overall and M Vesk (1994) Actin associated with plasmodesmata. Protoplasma 180: 169-184
- Corbesier, L., Vincent, C., Jang, S., Fornara, F., Fan, Q.; et al. . Science. 2007, 316 (5827): 1030–1033. PMID 17446353. doi:10.1126/science.1141752.
- Movement Protein of Tobacco Mosaic Virus Modifies Plasmodesmatal Size Exclusion Limit Science 20 October 1989: Vol. 246. no. 4928, pp. 377 - 379 SHMUEL WOLF, WILLIAM J. LUCAS, CARL M. DEOM,and ROGER N. BEACHY
- (PDF). [2013-08-12]. (原始内容 (PDF)存档于2010-02-16).