廣鹽性
廣鹽性,亦稱寬鹽性,是指一個物種(通常是魚類)能夠在鹽度大幅變化的水域生存的特征[1]。例如:黑花鱂能生存於淡水、半鹹水或鹹水水域之中[2],普通濱蟹则能存活於半鹹水和鹹水區域[3]。廣鹽性生物多棲息於河口灣[1]、潮池一類鹽度常出現變化的地方[4],而部分像鮭魚和鰻魚的物種,雖然本質上也屬廣鹽性生物[5],但在其生物生命週期中,牠們會按生活習性去變換生活環境;前者在產卵的時候游到淡水江河中產卵,產後返回海洋。與廣鹽性生物相反的物種稱為窄鹽性生物,意指牠們無法生存在鹽度浮動太大的水域[1][6]。牠們大多來自淡水區,轉往鹽水區生活便會死亡。同樣地,大部分海洋生物都是窄鹽性生物,轉到淡水生活也會死亡,是鹹淡水生物種無法超越的生理障礙[7]。
滲透壓調節

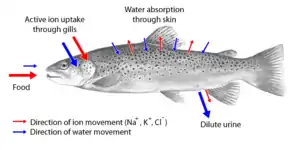
生物體面對不同環境變化如溫度變低、鹽度增加時,身體自然地因應情況來改變體內的有機/無機離子數量,從而調節細胞液泡的液體濃度,保持含水量,平衡體內外的滲透壓。滲透壓會讓細胞液維持適中狀態,既不會過於稀釋,又不會太濃。對於海洋無脊椎動物來說,即使海洋環境的離子組成有異,牠們可順應滲透壓去生存[8]。淡水魚有能力控制鹽分濃度[9][9],因其魚鰓上的線粒體細胞,即泌氯細胞阻擋鹽類滲入[10],魚繼而喝下海水,並通過低張性(鹽分被稀釋了)的尿液排出多餘的水分[9][11]。
海水魚的內部滲透壓較高,所以水分經滲透作用滲出體外,同時鹽分從外部滲入。魚鰓此時發揮功能,以減少過多鹽分進入魚的內部。另一方面,來自自然環境、高濃度的鈣有助淡水魚減低從魚鰓或魚體表面流失餘下的鹽分[9]。這樣不單降低對腎臟的負擔[9],也令血液裏的鹽分穩定下來[9]。相反,鹹水較體內液稠一些,因此鹹水魚會喝下大量海水[9],來補充在呼吸時失去的一部分體內水分[9]。
於海洋進行生物演化的魚類[1],包括鱈魚、吞拿魚等,多數是窄鹽性的[1];牠們用來呼吸的皮膚相對較薄[12],細胞由是缺少應付高滲透性分子的能力[1]。然而處於鹽度浮動大的水域中,有些種類展現出有效地調節滲透壓的能力,牠們就是廣鹽性生物。以鮭魚為例,鮭魚可棲息在兩個截然不同的環境:鹹水及淡水[13]。這是基於與生俱來的生活習性所致[14]。在海洋魚當中,鯊魚血液裏含有濃度相對較高的尿素[15]。尿素對活組織帶來傷害,為解決有關問題,鯊魚與一些魚種在身體內裏保留氧化三甲胺[16],用以對抗尿素毒性,溶質濃度達到1000以上毫滲透莫耳值(mOsm)[17]。還有,鯊魚將代謝後的氮化物,以尿素形式儲存於血液之中[18]。溶質濃度上升,血液濃度因而提高,滲透壓變得與海水相約,免卻吞水後再排鹽的生理機制[19]。
反觀淡水魚方面,對比海洋魚而言,大西洋魟含有濃度只有30至50個百分比的尿素和等壓滲透因子(osmolyte)[20]。儘管如此,體內液與外部水體兩者之間的滲透壓仍然令水分滲入牠們身體。牠們需排走比海洋物種還要多10倍的尿液,方可取得平衡[20]。
廣鹽性物種
- 廣鹽性魚類舉隅


 大西洋魟
大西洋魟 加州鱝
加州鱝 長吻魟[21]
長吻魟[21]
 大眼鯧
大眼鯧

 後鰭鋸鰩
後鰭鋸鰩
.jpg.webp)





潮間帶的鹽度水平變動大[7],雨水或河流淡水可導致低鹽度水平[8]。生存在這裏的河口灣魚種一定要是廣鹽性[4],能耐受廣闊的鹽度範圍[22]。個別生物體,像是飯蛸[8],水體中的鹽分就算明顯地被雨水沖淡或沖散都不足以令牠死亡,反之依然可生存至少數天[8]。此外,大西洋魟生理上是廣鹽性,卻棲息在淡水流域[23]。不像大西洋魟,沒有一個只活於淡水中的江魟科物種進化出專門的生理機制去調節滲透壓,可能礙於淡水拓殖出現在少於100萬年之內。不完整的基因進化亦是原因之一。
至於高鹽度水平,它發生在蒸發率高的地區[24],例如鹽沼和位於高位置的潮間帶岩池[1]。沼池被植物遮蔭有利蒸發率下降,紓緩鹽度給予的壓力。而且,鹽沼上的植物透過鹽腺排出鹽分,防止根部攝取過多分量。蘆葦、海三稜藨草等正是廣鹽性植物例子之一。
廣鹽性魚類遊走於鹹淡水之間生活,具有一定好處,諸如消除身上的寄生蟲[9]。由於鹽度水平迴異,牠們可選擇更多新食物[9][7],甚至避開獵食者視線[9],居於溫度穩定的棲息地[9]。
- 其他廣鹽性魚種例子



- 其他廣鹽性物種
.jpg.webp) 二藥藻
二藥藻_edit.jpg.webp) 澳洲斑點水母
澳洲斑點水母 鳥尾蛤
鳥尾蛤 紐西蘭泥蝸
紐西蘭泥蝸





參見條目
參考來源
- Smyth, K. & Elliott, M. (2016). Effects of changing salinity on the ecology of the marine environment. Maidenhead: Open University Press.
- Nordlie, F. G., Haney, D. C., & Walsh, S. J. (1992). Comparisons of salinity tolerances and osmotic regulatory capabilities in populations of sailfin molly (poecilia latipinna) from brackish and fresh waters. Copeia, 1992(3), 741–746.
- Josie Osborne. . Raincoast Education Society. 2009-01-01 [2021-12-04]. (原始内容存档于2021-12-04) (英语).
- Kling, L. J. & Jordaan, A. (Eds.). (2008). Inventory of tidepool and estuarine fishes in acadia national park. https://irma.nps.gov/DataStore/DownloadFile/474439 (页面存档备份,存于)
- Howard, J. (2018). Fish biology and fisheries. United Kingdom: Edtech Press.
- Rafferty, J. P. (Ed.). (2011). Biomes and ecosystems. New York City: Rosen Education Service .
- Kültz, D. (2015). Physiological mechanisms used by fish to cope with salinity stress. The Company of Biologists. 218, 1907-1914.
- Sakamoto, T., Ogawa, S., Nishiyama, Y., Akada, C., Takahashi, H., Watanabe, T., Minakata, H. & Sakamoto, H. (2015). Osmotic/ionic status of body fluids in the euryhaline cephalopod suggest possible parallel evolution of osmoregulation. Scientific Reports. 5.
- Wurts, W. A. (1998). Why can some fish live in freshwater, some in salt water, and some in both?. World Aquaculture. 29(1), 65.
- 許晉榮. . 水產試驗所: 32-36. 2013-06-01 [2021-12-06]. (原始内容存档于2021-12-06).
- Shu, Y., Lou, Q., Dai, Z., Dai, X., He, J., Hu, W. & Zhan, Y. (2016). The basal function of teleost prolactin as a key regulator on ion uptake identified with zebrafish knockout models. Scientific Reports. 6.
- Wells, J. (1996). Cutaneous respiration in octopus vulgaris. J Exp Biol 199, 2477–2483.
- . 世界自然基金會. 2015-03-19 [2021-12-04]. (原始内容存档于2021-12-04) (英语).
- Lennox, R. L., Pulg, U., Malley B., Gabrielsen, S. E., Hanssen, E. M., Cooke, S. J., Gauvin, K. B., Barlaup, B. T., & Vollset, K. W. (2020). The various ways that anadromous salmonids use lake habitats to complete their life history. Canadian Journal of Fisheries and Aquatic Sciences. 78(1), 90-100.
- Musick J. A. & Bonfil, R. (Eds.). (2005). Shark utilization. Rome: FAO.
- Summers, G., Wibisono,R. D., Hedderley, D. I., & Fletcher, G. C. (2016). Trimethylamine oxide content and spoilage potential of New Zealand commercial fish species. New Zealand Journal of Marine and Freshwater Research. 51(3), 393-405.
- Smith, S. L., Sim, R. B., & Flajnik, M. F. (Eds.). (2015). Immunobiology of the shark. Boca Raton: CRC Press.
- . CBC Radio. 2016-11-04 [2021-12-06]. (原始内容存档于2021-12-06) (英语).
- . American Museum of Natural History. [2021-12-06]. (原始内容存档于2022-01-23) (英语).
- Piermarini, P.M.; Evans, D.H. (PDF). Physiological and Biochemical Zoology. 1998, 71 (5): 553–560. PMID 9754532. doi:10.1086/515973. (原始内容 (PDF)存档于2020-07-31).
- Thorson, T. B. (1983). Observations on the morphology, ecology and life history of the euryhaline stingray, dasyatis guttata (bloch and schneider) 1801. Acta Biologica Venezuelica. 11(4), 95–126.
- . National Ocean Service. [2021-12-05]. (原始内容存档于2022-05-14) (英语).
- Janech, M.G., Fitzgibbon, W. R., Ploth, D. W., Lacy, E. R., & Miller, D.H. (2006). Effect of low environmental salinity on plasma composition and renal function of the Atlantic stingray, a euryhaline elasmobranch. Am J Physiol Renal Physiol. 291(4), 770-290.
- . National Weather Service. [2021-12-05]. (原始内容存档于2022-04-28) (英语).
- McCormick, S., Farrell, A. & Brauner, C. (Eds.). (2013). Fish physiology: Euryhaline fishes. Cambridge, Massachusetts: Academic Press.
- Palaima, A. (Ed.). (2012). Ecology, conservation, and restoration of tidal marshes: The san francisco estuary. Berkeley, California: University of California Press.