嘧啶代謝
嘧啶代謝是指一系列以嘧啶(衍生物)作為底物或代謝產物而進行的代謝途徑。在生物中自然存在的嘧啶環如組成嘧啶核苷酸的嘧啶鹼基,以及乳清酸(並非標準鹼基之一,但有著相近結構,因此可被修飾為鹼基之一的尿嘧啶)等,都在嘧啶代謝中有著重要的角色。
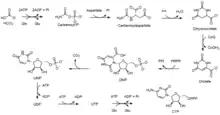
嘧啶核苷酸從頭合成 (合成代謝)
嘧啶在生物中會作為嘧啶核苷酸的一部分而被合成。此代謝途徑被稱為嘧啶核苷酸從頭合成(又稱乳清酸途徑),是生物合成嘧啶核苷酸的一個重要途徑。在此途徑中乳清酸(作為鹼基基礎)先被合成,再與核糖-5-磷酸以糖苷鍵鍵合而形成單磷酸乳清苷(OMP)。單磷酸乳清苷上乳清酸的部分其後被修飾為尿嘧啶,從而形成此途徑的第一個嘧啶核苷酸產物單磷酸尿苷(UMP)。單磷酸尿苷可進一步被磷酸化,再修飾其上的鹼基(尿嘧啶)而終被轉化為三磷酸胞苷(CTP)。[1] [2]
| 步驟 | 酶 | 進行地點(亦即酶的分佈) | 產物 | 說明[3] [4] |
|---|---|---|---|---|
| 1 | 氨甲醯磷酸合成酶 (CTPase) | (線粒體附近的)細胞質 | 氨甲醯磷酸 | 在此步驟穀氨醯胺先被水解而產生一個氨分子,氨分子再和碳酸氫以及兩個ATP反應,氨甲醯磷酸酯從而被合成。在動物細胞中,此步驟是此途徑的第一關鍵步驟,亦是此途徑的主要調節步驟。 2 ATP + 穀氨酰胺 + HCO3− + H2O → 2 ADP + Pi + 麩胺酸 + 氨甲醯磷酸酯 |
| 2 | 天門冬胺酸轉氨甲醯酶 (ATCase) | (線粒體附近的)細胞質 | 氨甲酰天冬氨酸 | 在此步驟氨甲醯磷酸酯上的磷酸基團被天冬氨酸取代。在細菌中此步驟是此途徑的第一關鍵步驟,同時亦是此途徑的主要調節步驟。 氨甲醯磷酸酯 + 天冬氨酸 → 氨甲醯天冬氨酸 + Pi |
| 3 | 二氫乳清酸酶 (DHOase) | (線粒體附近的)細胞質 | 4,5-二氫乳清酸 | 在此步驟氨甲醯天冬氨酸脫去一個水分子並環化成4,5-二氫乳清酸。此步驟是一個可逆反應。二氫乳清酸酶以鋅離子作為其輔因子。 氨甲醯天冬氨酸 + H+ → 4,5-二氫乳清酸 + H2O |
| 4 | 二氫乳清酸脫氫酶 (DODHase) | 線粒體(內膜表面上) | 乳清酸 (O) | 此步驟是此途徑唯一一個在線粒體進行的步驟。(此步驟的酶位於線粒體內膜表面上,因此上一步驟的產物4,5-二氫乳清酸需要從細胞質進入線粒體以進行此步驟)在此步驟4,5-二氫乳清酸脫去2個氫原子而被氧化成乳清酸,這些氫原子由輔酶Q接收,輔酶Q因而被還原並直接進入呼吸鏈。 4,5-二氫乳清酸 + CoQ (輔酶Q) → 乳清酸 + CoQH2 (還原態) |
| 5 | 乳清酸磷酸核糖轉移酶 (OPRTase) | (線粒體附近的)細胞質 | 單磷酸乳清苷 (OMP) | 上一個步驟的產物乳清酸重新離開線粒體。在此步驟磷酸核糖焦磷酸上的核糖-5-磷酸基團被轉移到乳清酸而形成單磷酸乳清苷。 乳清酸 + 磷酸核糖焦磷酸 (PRPP) → 單磷酸乳清苷 + PPi |
| 6 | 乳清酸核苷-5'-磷酸脫羧酶 (ODCase) | (線粒體附近的)細胞質 | 單磷酸尿苷 (UMP) | 在此步驟單磷酸乳清苷脫去二氧化碳(亦即脫羧反應)而成為單磷酸尿苷。 單磷酸乳清苷 → 單磷酸尿苷 + CO2 |
| \ | 單磷酸尿苷激酶 (UMPK) | 二磷酸尿苷 (UDP) | 在此步驟單磷酸尿苷被磷酸化。 單磷酸尿苷 + ATP → 二磷酸尿苷 + ADP | |
| \ | 核苷二磷酸激酶 (NDPK) | 三磷酸尿苷 (UTP) | 在此步驟二磷酸尿苷被磷酸化。 二磷酸尿苷 + ATP → 三磷酸尿苷 + ADP | |
| \ | 三磷酸胞苷合成酶 (CTPase) | 三磷酸胞苷 (CTP) | 在此步驟三磷酸尿苷上的尿嘧啶被磷酸化(從而被活化),其後再和一個氨分子(由穀氨酰胺水解而產生)反應而產生三磷酸胞苷。(尿嘧啶接上氨分子並脫去磷酸基團而成為胞嘧啶) 三磷酸尿苷 + ATP + 穀氨酰胺 → 三磷酸胞苷 + ADP + Pi + 麩胺酸 |
細節
值得注意的是,此途徑在動物和細菌中的進行有著一定的分別。
CAD蛋白
在細菌中,此途徑的所有六個步驟分別由六個獨立且受不同基因編碼的酶所催化。但在動物中,實際上首三個步驟的酶(CPSase,ATCase,DHOase)都受同一個基因編碼。它們和另一個酶(麩醯胺酸酶 GLNase)相結合而組成CAD蛋白(CAD protein)。
CAD蛋白是一個分子量為243kDa的多肽鏈,組成它的四個酶(GLNase,CPSase,ATCase,DHOase)分別成為了它的四個結構域(GLN 結構域,CPS 結構域,ATC 結構域,DHO結構域)。因此CAD蛋白是一個多功能酶而同時擁有以上四個酶的功能。所以在動物細胞中,更準確而言,是由CAD蛋白同時擔當起催化此途徑首三個步驟的角色(第一個步驟在GLN 和 CPS 結構域進行,第二和第三個步驟則分別在ATC 結構域和DHO結構域進行)。
另一方面,在細菌中所使用的CPSase和在動物中組成CAD蛋白的CPSase有著根本性的分別。細菌只有一種氨基甲醯磷酸合成酶(CPSase),它需要獨自提供所有代謝途徑所需的氨甲醯磷酸酯,因此在細菌中由CPSase催化產生的氨甲醯磷酸酯除了進入此途徑,亦會進入精氨酸合成途徑,所以在細菌中由CPSase催化的反應並非此途徑的第一關鍵步驟,而是其接下來由天冬氨酸氨基甲醯轉移酶(ATCase)所催化的反應(此步驟的產物則鎖定繼續此途徑)。而動物細胞則有兩種氨基甲醯磷酸合成酶,當中的 I 型(CPSase-I)專門負責尿素循環。而此途徑(第一個步驟)所使用的CPSase實際上是兩種氨基甲醯磷酸合成酶中的 II 型(CPSase-II),其合成的氨甲醯磷酸酯才可以(亦必須)供此途徑使用,所以在動物中由CPSase-II催化的步驟即是此途徑的第一關鍵步驟。總括而言,細菌中的CPSase並非專職於此途徑,而動物細胞中組成CAD蛋白的CPSase-II則是專職於此途徑。
[4] [5]
單磷酸尿苷合成酶
在動物中,此途徑另外兩個步驟-步驟五和六亦有著類似的情況。這兩個步驟所使用的酶(OPRTase,ODCase)亦受同一個基因編碼。它們相結合而組成單磷酸尿苷合成酶(UMPS)。
單磷酸尿苷合成酶的分子量為52kDa,組成它的兩個酶(OPRTase,ODCase)分別成為了它的兩個結構域(OPRT 結構域,ODC 結構域)。和CAD蛋白一樣,單磷酸尿苷合成酶是一個多功能酶並同時擁有以上兩個酶的功能。因此在動物中(更準確而言)是由單磷酸尿苷合成酶同時擔當起催化此途徑第五,第六個步驟的角色(第五個步驟在OPRT 結構域進行,第六個步驟則在ODC 結構結構域進行)。
研究指出動物細胞選擇使用多功能的單磷酸尿苷合成酶,而非獨立分開的 OPRTase 和 ODCase 的原因是由於單磷酸尿苷合成酶的兩個結構域之間的共價鏈結令其穩定性比起單體形式的 OPRTase 和 ODCase 更高,因此其催化能力亦相對有所提升。所以在動物細胞中所需的單磷酸尿苷合成酶的濃度則比起細菌中所需的OPRTase 和 ODCase 的濃度明顯地低。
[4] [6]
氨基甲醯磷酸合成酶 II
在動物中,由氨基甲醯磷酸合成酶 II (作為CAD蛋白的一部分)催化的步驟是此途徑最主要的調節步驟。氨甲醯磷酸合成酶 II 以異位調節的方式受磷酸核糖焦磷酸(為此途徑第五個步驟的底物之一)活化而受三磷酸尿苷(為此途徑其中的一個產物)反饋抑制,從而調節總反應速率。
天門冬胺酸轉氨甲醯酶
在細菌中,由天門冬胺酸轉氨甲醯酶 (作為獨立的酶)催化的步驟則是此途徑最主要的調節步驟。天門冬胺酸轉氨甲醯酶以異位調節的方式受三磷酸尿苷和三磷酸胞苷(為此途徑的最終產物)反饋抑制,從而調節總反應速率。然而在動物中,天門冬胺酸轉氨甲醯酶對於此途徑並沒有調節作用(不受調節)。
嘧啶還原途徑 (分解代謝)
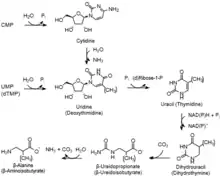
在生物中所存在的自由嘧啶環有一大部分來自於被分解的嘧啶核苷酸(嘧啶核苷酸先被分解為相應的核苷,這些核苷再被進一步分解成嘧啶鹼基),這些嘧啶環會通過嘧啶還原途徑而被降解。視乎被分解的嘧啶環是尿嘧啶或胸腺嘧啶,它們會在一連串的反應後分別被降解而轉化為β-丙胺酸或β-氨基異丁酸鹽。
單磷酸胞苷 (CMP) 和 單磷酸尿苷 (UMP) 在此途徑中會先被轉化為尿嘧啶,因而終會被轉化為β-丙胺酸。而去氧單磷酸胸苷 (dTMP) 在此途徑中則會先被轉化為胸腺嘧啶,因此終會被轉化為β-氨基異丁酸鹽。在此途徑中所產生的副產物(氨和二氧化碳)則會進入尿素循環。[3] [8]
| 步驟 | 酶 | 產物 | 說明 |
|---|---|---|---|
| (單磷酸胞苷版本) | |||
| (1) | 5'-核苷酸酶 | 胞苷 (C) | 單磷酸胞苷 + H2O → 胞苷 + Pi |
| (2) | 胞苷脫氨酶 | 尿苷 (U) | 胞苷 + H2O → 尿苷 + NH3 |
| 單磷酸尿苷版本 | |||
| 1 | 5'-核苷酸酶 | 尿苷 (U) | 單磷酸尿苷 + H2O → 尿苷 + Pi |
| 2 (3) | 尿苷磷解酶 | 尿嘧啶 | 尿苷 + Pi → 尿嘧啶 + 核糖-1-磷酸 |
| 3 (4) | 二氫嘧啶脫氫酶 | 二氫尿嘧啶 | 尿嘧啶 + NADH/NADPH + H+ → 二氫尿嘧啶 + NAD+/NADP+ |
| 4 (5) | 二氫嘧啶酶 | β-脲基丙酸鹽 | 二氫尿嘧啶 + H2O → β-脲基丙酸鹽 |
| 5 (6) | β-脲基丙酸酶 | β-丙胺酸 | β-脲基丙酸鹽 + H2O → β-丙胺酸 + NH3 + CO2 |
| 步驟 | 酶 | 產物 | 說明 |
|---|---|---|---|
| 去氧單磷酸胸苷版本 | |||
| 1 | 5'-核苷酸酶 | 去氧胸苷 (dT) | 去氧單磷酸胸苷 + H2O → 去氧胸苷 + Pi |
| 2 | 去氧胸苷磷解酶 | 胸腺嘧啶 | 去氧胸苷 + Pi → 胸腺嘧啶 + 去氧核糖-1-磷酸 |
| 3 | 二氫嘧啶脫氫酶 | 二氫胸腺嘧啶 | 胸腺嘧啶 + NADH/NADPH + H+ → 二氫胸腺嘧啶 + NAD+/NADP+ |
| 4 | 二氫嘧啶酶 | β-脲基異丁酸鹽 | 二氫胸腺嘧啶 + H2O → β-脲基異丁酸鹽 |
| 5 | β-脲基丙酸酶 | β-氨基異丁酸鹽 | β-脲異丁酸鹽 + H2O → β-氨基異丁酸鹽 + NH3 + CO2 |
產物
嘧啶還原途徑是β-丙胺酸和β-氨基異丁酸鹽在生物中的重要來源之一。這兩種產物都會進入而參與其他的代謝途徑。
嘧啶核苷酸補救合成 (合成代謝)
嘧啶核苷酸被分解後而衍生出的核苷和鹼基並不一定進入嘧啶還原途徑以被繼續分解,實際上它們亦可被重新利用來合成新的嘧啶核苷酸。此途徑被稱為嘧啶核苷酸補救合成,所耗用的能量相對於從頭合成途徑較少。對於部分無法進行從頭合成途徑的生物,此補救合成途徑尤其重要。而此途徑在植物的葉綠體中亦特別活躍。
多種的嘧啶核苷(尿苷 U、胞苷 C、去氧胞苷 dC、胸苷 T) 都會被此途徑用作合成相應的核糖核苷酸或去氧核糖核苷酸。在各種的鹼基之中,則只有尿嘧啶被此途徑使用。[11]
參考資料
- Soderberg, Timothy. . : P. 72–74 [2021-01-27]. (原始内容存档于2018-04-18).
- W H Freeman. . : Section 25.1 [2021-01-27]. (原始内容存档于2019-12-16).
- Barbara A., Moffat; Hiroshi, Ashihara. . The Arabidopsis Book, 2002(1). 4 April 2002 [2021-01-27]. doi:10.1199/tab.0018. (原始内容存档于2021-02-26).
- David R., Evans; Hedeel I., Guy. . Minireviews Enzymology. 6 August 2004. doi:10.1074/jbc.R400007200.
- María, Moreno-Morcillo; Araceli, Grande-García 等. . Structure. 6 June 2017 [2021-01-27]. doi:10.1016/j.str.2017.04.012. (原始内容存档于2021-02-05).
- Michael J., Yablonski; Daniel A., Pasek 等. . Enzymology. 3 May 1996. doi:10.1074/jbc.271.18.10704.
- Andrew N, Lane; Teresa W-M, Fan 等. . Nucleic Acids Research. 27 February 2015 [2021-01-27]. doi:10.1093/nar/gkv047. (原始内容存档于2021-01-20).
- Jinyu, Yin; Yifeng, Wei 等. . Microbiology. 25 Oct 2019. doi:10.1074/jbc.RA119.010406.
- Anutthaman Parthasarathy, Michael A. Savka, André O. Hudson. . Front Plant Sci. 18 Jul 2019 [2021-05-31]. doi:10.3389/fpls.2019.00921. (原始内容存档于2022-07-06).
- Shi, CX., Zhao, MX., Shu, XD. 等. . Sci Rep. 24 Feb 2016 [2021-05-31]. doi:10.1038/srep21924. (原始内容存档于2022-05-16).
- Christopher Leija, Filipa Rijo-Ferreira, Lisa N. Kinch, 等. . PLoS Pathog. 11 Nov 2016. doi:10.1371/journal.ppat.1006010.
外部連結
- (英文)嘧啶生物從頭合成概觀 (页面存档备份,存于)
- (英文)嘧啶生物從頭合成筆記 (页面存档备份,存于)