星宿龙属
星宿龙属(属名:Xingxiulong,意为“星宿桥之龙”)是中国早侏罗世两足蜥脚形类的一属,仅含一个物种,即程氏星宿龙(X. chengi),由王娅明等人叙述于2017年。已知化石包括三件标本,其中两件为成体,另一件为发育未完全的个体,它们埋藏于同一地点且互相补充,构成了该属的大部分骨骼。据估计,本属成体体长约为4—5(13—16英尺),高约1—1.5(3英尺3英寸—4英尺11英寸)。系统发育分析表明,星宿龙与同时期的金山龙息息相关,尽管其在蜥脚形类和大椎龙科之外的其他位置也是合理的。
| 星宿龙属 化石时期: | |
|---|---|
 | |
| 星宿龙骨骼组图(a~n)与骨骼重建(o) | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 演化支: | †悍龍類 Bagualosauria |
| 演化支: | †板龙类 Plateosauria |
| 演化支: | †大脚类 Massopoda |
| 演化支: | †蜥脚型类 Sauropodiformes |
| 属: | †星宿龙属 Xingxiulong Wang et al., 2017 |
| 模式種 | |
| †程氏星宿龙 Xingxiulong chengi Wang et al., 2017 | |
尽管二者有着密切联系,但星宿龙与金山龙(以及大多数基底蜥脚形亚目动物)的显著不同之处在于前者具有许多类似蜥脚亚目的特征,包括:骶骨具有四块椎骨;耻骨顶部特别长;股骨、脚部的第一、第五跖骨及肩胛骨宽而结实。这些特征可能代表了其对支持巨大体重特别是大型肠道的适应性。但与蜥脚类不同的是星宿龙仍为双足动物。
叙述

星宿龙是一种中等大小的蜥脚形类,成年后体长可达4—5(13—16英尺),臀高为1—1.5(3英尺3英寸—4英尺11英寸)。从颅骨化石完全融合来看,该属两个较大的标本LFGT-D0002和LFGT-D0003是成年个体;第三个标本LFGT-D0001相对而言小了14%,由于椎骨融合不完全而被认为是一具亚成体。[1]
颅骨与颌骨
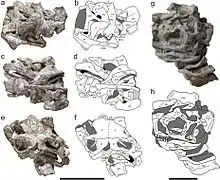
与同时期的禄丰龙不同,[2]星宿龙的上颌骨侧面没有嵴,保留了11个牙槽。再往后方,在眼窝前面,泪骨在其前端顶部附近有一个突出的突起,该特征也可见于禄丰龙、[2]远食龙、大椎龙[3]和里奥哈龙身上,但不包括云南龙、金山龙或随后的蜥脚形类。颧骨与眶后骨之间的接触面相当长,类似禄丰龙,而非云南龙。[4]在颧骨的三个分支中,向后与向上突出的两个分支成80°角,类似板龙和槽齿龙,但这一角度比其它蜥脚形亚目大得多。[1][4][5][6]
在颅骨底部,方轭骨有两个分支,一个指向前方,另一个指向上方。它们彼此大致垂直,与禄丰龙(45度角)、云南龙(60度角)和金山龙(110度角)不同。在方轭骨上方,方骨具有两个关节髁突(articulating condyles),其中一个呈三角形,朝外;另一个更圆的朝内。后者更靠近方骨底部,类似陆丰龙和云南龙,但有别于板龙。在颅骨后部,顶骨与枕骨间有一个突出的后顶窗(postparietal fenestra)。枕骨上端的底部向前倾斜,以使颅骨的底部变圆。基蝶状骨横突(basipterygoid processes)细长,像板龙一样向下且向外突出,彼此成80°角,与禄丰龙和金山龙不同。[1]
与禄丰龙、云南龙和金山龙相比,星宿龙的下颌窗(mandibular fenestra)的隅骨和上隅骨延伸得更远,更接近于远食龙和板龙。[7]其关节带有向内伸出的锥状突(pyramidal process)作为下颌关节的延伸;在其后端,它还具有一个向上延伸的凸舌状突起,该特征在科罗拉多斯龙、[8]金山龙和一种迄今尚未命名的蜥脚形亚目身上也能看见。[1]
椎骨
星宿龙的颈部有十块颈椎。前寰椎(proatlas)是位于寰椎前方的一块萎缩性椎骨,在其前方被枕骨大孔(foramen magnum)的顶边界定。尽管由于化石损坏而很难叙述寰椎本身,但是紧随其后的轴具有相对较短的中心,侧面和底部略微受压。总的来说,剩余颈椎相对较短,只有它们长的2.5到3倍,像禄丰龙一样愈向后愈短;而其它基底蜥脚形亚目[9][10](包括金山龙)的颈椎长度通常大于其高度3至4倍。第4至9节颈椎的中央底部有突出的龙骨,形如禄丰龙、金山龙和其他基底蜥脚形亚目恐龙。在颈后部附近,神经棘变得略呈板状。[1]
星宿龙的十四节背椎略呈双凹型(amphicoelous)或两端凹陷,属于典型的基底蜥脚形亚目特征。其同样典型的特征是第一至第三背椎下侧有叶片状龙骨,这是其它恐龙所没有的。此外,身体前部附近的背椎具有典型的低、短且略呈板状的神经棘。然而,不同寻常的是在后三个背椎上也可见到这类神经棘,而只有在像艾雷拉龙和始盗龙这样的基底蜥臀目动物中才具备该特征。在中后背椎上,神经棘后上角向外突出以产生凹形后缘,此特征可见于其它一些基础蜥脚类身上,而在禄丰龙、金山龙、云南龙和里奥哈龙身上则没有看到。[1]
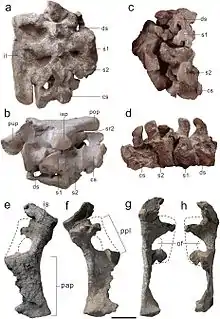
在基础蜥脚形亚目中,星宿龙有四节骶椎。这是一种典型特征,通常出现在更多的蜥脚形亚目身上,如黑丘龙、利奥尼拉龙[11]巨脚龙[12]和蜀龙。这些骶椎中的第一节,即背骶(dorsosacral)与下一节骶骨的融合不好;而在横突(transverse processes)上,它也没有完全融合到相应的肋骨。背骶和第一对骶肋共同向前延伸以接触髂骨。第二和第三骶椎(两个“原始骶骨”)长度大致相同且彼此融合良好。正像其他基底蜥脚形类一样,第三骶椎横突也向后向外突出;第四骶椎的横突和肋同样如此,它们在后侧再次接触髂骨。所有神经棘皆呈板状,就像最后几节背椎上的一样。[1]
星宿龙尾部保留的尾椎可能超过35个。这些椎骨总体较高,非常坚固,并且具有像其它基底蜥脚形亚目一样的凹面,它们也似乎皆呈两凹型。前几个尾椎的横突宽而平坦,并向上向外突出;位于更后方的尾椎横向突更加细长和水平,神经棘高而细长,并有些向后突出。[1]
四肢
星宿龙的肩胛骨两端均已扩大,底端宽度为其长度的56%,顶端宽度为其长度的49%。其它各种基础蜥脚形类动物(包括禄丰龙和金山龙)的肩胛骨底端更为膨大,但一般较为细长。相反,雷前龙[13]和莱森龙具有类似的健壮的肩胛骨,但是这两个分类单元的肩胛骨顶端更加扩展。与金山龙一样,其肩胛骨轴的最大宽度为整件骨骼长度的19%至20%。雷前龙[13]和莱森龙的轴更为坚固,而大多数基底蜥脚形亚目的轴则更窄。[1]
与大多数基底蜥脚形亚目(包括禄丰龙和云南龙)相反,星宿龙肱骨顶端内表面的结节(tuberosity)发育较差。像云南龙和金山龙一样,其尺骨长度大约是肱骨的61%。而在禄丰龙身上,该比例为68%,导致尺骨更长。尺骨的顶端相当大,有突出的前内侧突(anteromedial process)和前外侧突(anterolateral process)。这些突起连同较浅的桡窝(radial fossa)共同形成了尺骨与桡骨的关节。后一骨骼很细长,长度约为肱骨的54%。星宿龙手部至少有四根手指的化石是已知的,但没有得到很好的叙述。[1]
总的来说,髂骨与其它基底蜥脚形亚目相似,前部不超过耻骨柄(pubic peduncle)或其与耻骨连接部分的水平面。与其它基底蜥脚形亚目动物不同,其后端略呈方形而不尖锐,从侧面看时底部很凹(在其它基底蜥脚形类动物中,它大部分是直的,甚至是凸的)。坐骨柄(ischial peduncle)或与坐骨的连接部分在其底端有一个小的、突出的突起。耻骨顶端(称为耻骨板)相对较长,占骨头长度的40%,而底部(称为耻骨领)则相对较短。这不同于其它蜥脚形亚目,但类似于一些基础蜥脚类。耻骨领(pubic apron)外表面有些凹陷,底端向前、后扩展到整个骨骼长度的16%。最后,坐骨膨胀的闭孔板(obturator plate)在其侧面上有一个凹槽。[1]
与云南龙和金山龙不同,在股骨上,小转子(lesser trochanter)向股骨头下方延伸。从正面看,转子与其它除雷前龙和黑丘龙(它们更靠近外边缘)以外的基础蜥脚形亚目一样靠近骨骼中线。同样,与许多基础蜥脚形亚目[8](包括禄丰龙)相反,第四转子(fourth trochanter)位于中线附近。在胫骨底部的两个突起中,后一个较薄,并且比前一个更加向外突出,而非向下。这与大多数其它蜥脚形亚目动物不同,后者两个突起的厚度均等或者均等地向外突出(如陆丰龙和金山龙),抑或前者更向外突出(如云南龙)。[1]
距骨背部靠近底端的位置有一个突起。在鼠龙[6]化石上也可见此类突起,尽管后者发育更好且更靠近中线。在跖骨中,第一个是最宽和最坚固的。第五跖骨顶端异常膨胀,其宽度为整件骨骼长度的85%。在其它蜥脚形类中,这个数字通常为50%至77%,[14]尽管雷前龙也可能具有类似扩张的跖骨。在跖骨下方,五根脚趾的指式(phalangeal formula)为2-3-4-5-1。[1]
发现与命名
LFGT-D0002为星宿龙的正模标本,来自一只成熟个体,包括部分颅骨和颌骨;第七至第九节颈椎、第八至第十四节背椎、整个骶骨和三十五节尾椎;肋骨和人字骨碎片;左髂骨以及部分耻骨和坐骨;股骨、胫骨和部分腓骨、左踝骨和双脚(几乎完整)。[1]
该属存在两个参考标本。LFGT-D0003也是一只成熟个体,包括部分颅骨和颌骨;第三至第十节颈椎以及所有的背骨和大部分骶骨;肋骨和人字骨碎片;肩胛骨、肱骨、尺骨和桡骨以及手的一部分;部分髂骨(右侧完整)和耻骨(左侧完整);左股骨、部分胫骨和腓骨(右侧完整),以及右踝骨和部分脚。LFGT-D0001较小,可能发育未成熟,由骨轴(axis)、第三至第十节颈椎、全部背椎骨和骶骨以及第十九节尾椎;肋骨和人字骨碎片;右肩胛骨和髂骨,以及部分右侧耻骨和坐骨;部分股骨和左胫骨以及左踝骨。[1]
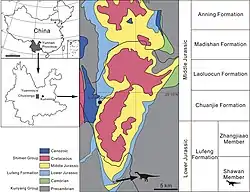
这些标本2013年发现于中国云南省禄丰县三棵树村附近且埋葬在一起,目前保存在禄丰县国土资源局。保存标本的岩石为紫色粉砂质泥岩,属于早侏罗世禄丰组(Lufeng Formation)沙湾段(Shawan Member ),年代初步确定为赫唐阶。[1]
2017年,王娅明、尤海鲁和王涛对星宿龙进行叙述。属名指建于中国明朝的星宿桥(字面意思是“星座”)。同时,种名chengi纪念程政武教授,他为包括禄丰地区在内的中国生物地层学(biostratigraphy)研究做出了重大贡献,并于2015年去世。[1]
分类学
基于B·W·麦克菲等人的数据集进行的2017年系统发育分析(发表于2015年,叙述了新属造雨龙[15])发现,一旦从数据集中移除贝里肯龙,星宿龙便是蜥脚形类的基础成员。在该分类群中,它最接近同时期的金山龙。星宿龙与金山龙共有的特征包括:颞下窗(infratemporal fenestra)完全置于眼窝后面;肩胛骨的宽度至少是其长度的20%;耻骨领(pubic apron)或耻骨底部具有凹入的外表面;耻骨领底部的膨胀至少是整个骨骼长度的15%; 股骨头(femoral head)与股骨干(femoral shaft)的横截面轴线之间的角度约为30°。通过此分析恢复的拓扑结构(topology)如下所示。[1]
| 大脚类 Massopoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
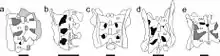
测试了星宿龙的各种其他系统发育位置;其中,最合理的选择是将其作为大脚类的基础成员放置在大椎龙科科或蜥脚形类之外,这仅需要两个额外的进化步骤即可。这可能是由于眶后骨弯曲的上缘和上颌关节后方存在突起的原因,与大椎龙科或蜥脚形类相比,这些特征可以认为是相对原始的。这种替代方案将在此进化等级中涉及大量镶嵌进化(mosaic evolution)。需要另外三个进化步骤的位置将星宿龙放在金山龙和近蜥龙之间或云南龙和金山龙之间。[1]
奇怪的是,星宿龙具有许多通常可在蜥脚类身上见到的特征,包括:具有四块椎骨的骶骨(鼠龙也可以通过趋同进化获得);[6][11][16]长的耻骨板(pubic plate)或耻骨顶部占骨长的40%(此数字在大多数其他基底蜥脚形亚目中占25%,[8]在大多数蜥脚类中占33%,[17]在圆顶龙形类中占45%至50%[18]);相对较为坚固的股骨干、[11]第一节跖骨、[19]第十五节跖骨顶端以及肩胛骨。这些独特的特征可能是趋同进化而不是具有共同祖先的产物。[1]
古生物学
星宿龙骨骼的整体坚固性,尤其是臀部、股骨和脚,都与蜥脚类趋同,这表明其内脏相对较大,整体上体重较高。然而,与蜥脚类不同,星宿龙是两足动物(bipedal);它缺乏蜥脚类动物对四足(quadrupedal)行走的适应能力,包括相对较长的前肢和尺骨侧面前方有突出的突起,而股骨干相对笔直,其尺骨和股骨总的来说更像是典型的蜥脚形亚目。星宿龙、金山龙和云南龙的肩胛骨大而强壮,可能在其两足行动时增加了前肢的活动性,但这一特性后来被视作蜥脚类对四足行走的一种适应性。[1][15]
古生态学
禄丰组(Lufeng Formation)含有湖泊、河流和漫滩沉积物中的泥岩和粉砂岩(siltstone)。[20]所知于禄丰组的许多蜥脚形亚目包括许氏禄丰龙(Lufengosaurus huenei)、巨型禄丰龙(L. magnus)、黄氏云南龙(Yunnanosaurus huangi)、中国兀龙("Gyposaurus" sinensis)、新洼金山龙(Jingshanosaurus xinwaensis)、禄丰楚雄龙(Chuxiongosaurus lufengensis)、[1]孙氏细细坡龙(Xixiposaurus suni)、孙氏益州龙("Yizhousaurus sunae")[21]以及不完整硕鳄(Pachysuchus imperfectus)。此外,还有兽脚类的三叠中国龙(Sinosaurus triassicus)、尹氏芦沟龙(Lukousaurus yini)、[20]金时代龙(Shidaisaurus jinae)[21][22]以及出口峨山龙(Eshanosaurus deguchiianus);[23]鸟臀目的奥氏大地龙(Tatisaurus oehleri)[20]和禄丰卞氏龙(Bienosaurus lufengensis);鳄形超目的敏捷裂头鳄(Dibothrosuchus elaphros)、许氏扁颌鳄(Platyognathus hsui)、甲板小鳄(Microchampsa scutata)和张家洼滇鳄(Dianosuchus changchiawaensis)以及禄丰滇中龙(Dianchungosaurus lufengensis);分类不明的古龙类曲条鳄(Strigosuchus licinus);喙头蜥目的佩氏克利沃蜥(Clevosaurus petilus)、王氏克利沃蜥(C. wangi)和麦氏克利沃蜥(C. mcgilli);三列齿兽科的二齿兽云南卞氏兽(Bienotherium yunnanense)、小卞氏兽(B. minor)和巨型卞氏兽(B. magnum)、精致禄丰兽(Lufengia deltcata)、短吻云南齿兽(Yunnanodon brevirostre)以及长吻滇中兽(Dianzhongia longirostrata);哺乳类的瑞氏中华锥齿兽(Sinoconodon rigneyi)、奥氏摩根齿兽(Morganucodon oehleri)、黑果蓬摩根齿兽(M. heikoupengensis)以及小昆明兽(Kunminia minima);一种原颚龟科的乌龟;以及一种“迷齿亚纲”的两栖类。[20]
参见
- 2017年初龙类研究
参考资料
- Wang, Y.-M.; You, H.-L.; Wang, T. . Scientific Reports. 2017, 73: 41881. PMC 5312170
 . PMID 28205592. doi:10.1038/srep41881.
. PMID 28205592. doi:10.1038/srep41881. - Barrett, P.M.; Upchurch, P.; Wang, X.-L. . Journal of Vertebrate Paleontology. 2005, 25 (4): 806–822. JSTOR 4524508. doi:10.1671/0272-4634(2005)025[0806:COOLHY]2.0.CO;2.
- Sues, H.-D.; Reisz, R.R.; Hinic, S.; Raath, M.A. . Annals of Carnegie Museum. 2004, 73 (4): 239–257.
- Barrett, P.M.; Upchurch, P.; Zhou, X.-D.; Wang, X.-L. . Zoological Journal of the Linnean Society. 2007, 150 (2): 319–341. doi:10.1111/j.1096-3642.2007.00290.x.
- Lu, J.; Kobayashi, Y.; Li, T.; Zhong, S. . Acta Geologica Sinica. 2010, 84 (6): 1336–1342. doi:10.1111/j.1755-6724.2010.00332.x.
- Otero, A.; Pol, D. . Journal of Vertebrate Paleontology. 2013, 33 (5): 1138–1168. doi:10.1080/02724634.2013.769444.
- Prieto-Marquez, A.; Norell, M. . American Museum Novitates. 2011, 3727 (3727): 1–58. doi:10.1206/3727.2. hdl:2246/6138.
- Apaldetti, C.; Martinez, R.N.; Pol, D.; Souter, T. . Journal of Vertebrate Paleontology. 2014, 34 (5): 1113–1132. doi:10.1080/02724634.2014.859147.
- Martinez, R.N. . Journal of Vertebrate Paleontology. 2009, 29 (1): 142–164. doi:10.1671/039.029.0102.
- Apaldetti, C.; Martinez, R.N.; Alcober, O.A.; Pol, D. . PLOS One. 2011, 6 (11): e26964. PMC 3212523 . PMID 22096511. doi:10.1371/journal.pone.0026964.
- Pol, D.; Garrido, A.; Cerda, I.A. . PLOS One. 2011, 6 (1): e14572. PMC 3027623 . PMID 21298087. doi:10.1371/journal.pone.0014572.
- Jain, S.L.; Kutty, T.S.; Roy-Chowdhury, T.; Chatterjee, S. . Proceedings of the Royal Society B: Biological Sciences. 1975, 188 (1091): 221–228. doi:10.1098/rspb.1975.0014.
- McPhee, B.W.; Yates, A.M.; Choiniere, J.N.; Abdala, F. . Zoological Journal of the Linnean Society. 2014, 171 (1): 151–205. doi:10.1111/zoj.12127.
- McPhee, B.W.; Choiniere, J.N.; Yates, A.M.; Viglietti, P.A. . Journal of Vertebrate Paleontology. 2015, 35 (5): e980504. doi:10.1080/02724634.2015.980504.
- McPhee, B.W.; Bonnan, M.F.; Yates, A.M.; Neveling, J.; Choiniere, J.N. . Scientific Reports. 2015, 5: 13224. PMC 4541066 . PMID 26288028. doi:10.1038/srep13224.
- Upchurch, P. . Zoological Journal of the Linnean Society. 1998, 124 (1): 43–103. doi:10.1006/zjls.1997.0138.
- Wilson, J.A.; Sereno, P.C. . Journal of Vertebrate Paleontology. 1998, 18 (sup002): 1–79. doi:10.1080/02724634.1998.10011115.
- Salgado, L.; Coria, R.A.; Calvo, J.O. . Ameghiniana. 1997, 34 (1): 3–32. ISSN 0002-7014.
- Yates, A.M.; Bonnan, M.F.; Neveling, J.; Chinsamy, A.; Blackbeard, M.G. . Proceedings of the Royal Society B: Biological Sciences. 2009, 277 (1682): 787–794. PMC 2842739 . PMID 19906674. doi:10.1098/rspb.2009.1440.
- Luo, Z.; Wu, X.-C. . Fraser, N.C.; Sues, H.-D. (编). . Cambridge: Cambridge University Press. 1997: 251–270 [2020-12-10]. ISBN 9780521458993. (原始内容存档于2018-10-14).
- Xing, L.; Rothschild, B.M.; Ran, H.; Miyashita, T.; Persons, W.S.; Sekiya, T.; Zhang, J.; Wang, T.; Dong, Z. . Acta Palaeontologica Polonica. 2015, 60 (3): 643–649. doi:10.4202/app.00001.2013.
- Wu, X.-C.; Currie, P.J.; Dong, Z.; Pan, S.; Wang, T. . Acta Geologica Sinica. 2009, 83 (1): 9–24. doi:10.1111/j.1755-6724.2009.00002.x.
- Xu, X.; Zhao, X.; Clark, J.M. . Journal of Vertebrate Paleontology. 2001, 21 (3): 477–483. JSTOR 20061976. doi:10.1671/0272-4634(2001)021[0477:ANTFTL]2.0.CO;2.