大角鹿
大角鹿(學名:[1]),又名巨大角鹿、巨型鹿或愛爾蘭駝鹿[2],是已發現體型最大的鹿,生存於更新世中期及全新世中期的歐亞大陸,由愛爾蘭至西伯利亞的貝加爾湖。最近年代的化石發現於西伯利亞,由放射性碳定年法測定後為屬於7700年前[3][4]。大角鹿的大量化石現正在愛爾蘭都柏林的愛爾蘭自然歷史博物館展覽。
| 大角鹿 | |
|---|---|
 | |
絕滅(約公元前5700年) | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 哺乳纲 Mammalia |
| 目: | 偶蹄目 Artiodactyla |
| 科: | 鹿科 Cervidae |
| 属: | †大角鹿屬 Megaloceros |
| 种: | †大角鹿 M. giganteus |
| 二名法 | |
| †Megaloceros giganteus Blumenbach, 1799 | |
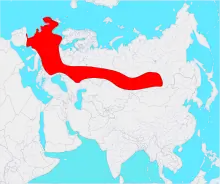 | |
| 晚更新世時期大角鹿的分布區域 | |
| 異名 | |
| |
其种加词“Giganteus”意为“巨大的”。
傳統上大角鹿被稱為愛爾蘭駝鹿。雖然有大量的骨骼在愛爾蘭的酸沼發現,但牠們卻不只限於愛爾蘭,加上牠其實與駝鹿和加拿大馬鹿不是近親,故現時很多學者都只稱呼牠為大角鹿。[5][3][6]現今的物種都可能是從M. antecedens在近埃姆間冰期演化而來。M. antecedens有時被認為是一個亞種,稱為M. giganteus antecedens,牠的鹿角相似但更為複雜。
雖然有一項研究指出大角鹿現存的最近親為歐洲馬鹿[7],但是其他系統分類學上的研究則多認為大角鹿是黇鹿的姊妹演化支[6][8][9][10]。
分類學
研究歷史

大角鹿最初由愛爾蘭醫師湯瑪斯·莫利紐茲根據於都柏林達迪斯敦發現的巨大鹿角在1956年進行正式發表,在當時他認為這鹿角為駝鹿的鹿角,並認為這代表駝鹿曾廣泛分布於愛爾蘭島上[11]。法國科學家喬治·居維葉為第一個挑戰這項觀點的人,他認為這些鹿角並不屬於任何已知現存的物種。
鹿角

在20世紀之前,大角鹿的巨大鹿角曾被用來否定達爾文的演化論,認為這麼大的角顯然沒有好處,是源自定向演化,鹿角會不斷的增大,繼而由於過份的大以致大角鹿不能正常覓食或是鹿角與樹枝糾纏進而滅絕[6]。另一些人則認為鹿角持續受到強烈的性選擇,因雄鹿需要打鬥才能得到雌鹿[12]。史蒂芬·古爾德於1974年指相對於一般的鹿,大角鹿不單體型上較大,鹿角在比例上亦較為大,顯示在發育期間的身體及鹿角的生長率不同[5]。大角鹿的體型剛好適合較大的角,故大角鹿的祖先早已有足夠大的角來演化。而古爾德更指鹿角的大小及位置正是由性選擇來維持:這對鹿角在形態上很不適合打鬥,但其在頭顱骨的位置則在攻擊敵人及吸引異性方面十分完美。大角鹿不像其他的鹿,牠們根本不用轉動頭部來顯示其角的美態,單單從其正面就可以看到。
演化

大角鹿屬於大角鹿屬,該屬最古老的物種可以追溯至更新世早期[13]。M. giganteus被認為並非是更新世中期常見於歐洲的中等體型物種M. savini的直屬後代。於更新世早期(約120萬年前)希臘所發現的未命名大角鹿屬物種化石被認為與大角鹿親緣關係較M. savini近,因為兩者第四下前臼齒(P4)齒冠型態結構十分類似[14]。大角鹿最古老的化石出土自諾福克郡的霍默斯菲爾德,地層年代約為450,000年前;以及德國的穆爾河畔施泰因海姆(化石被認為屬於亞種M. g. antecedens),地層年代約為400,000–300,000年前 [15]。
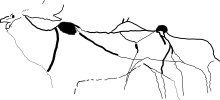
大部分已知的大角鹿遺骸發現於更新世晚期,超過一百頭的大角鹿化石是在都柏林巴利貝格的酸沼之中發現[17]。
大角鹿屬於可能為多系群的大角鹿族,其他屬於大角鹿族的演化支包括Megaceroides、Praemegaceros、真枝角鹿及腫骨鹿。

2005年,自4根鹿角與骨頭中的細胞色素b抽取到了兩段粒線體DNA,分析這兩段粒線體DNA後發現,大角鹿可能曾與歐洲馬鹿進行雜交,這在現存鹿物種之中並不少見。由於都具有掌狀的鹿角,大角鹿被認為與黇鹿為近親,然而粒線體DNA的分析結果卻認為大角鹿可能與鹿屬的關係較近[18]。然而,2015年接續粒線體DNA分析的研究結果卻反駁了2005年的研究結果,並證明大角鹿確實與黇鹿的親緣關係較近[9],此外兩者的骨迷路型態也十分近似,這代表著2005年的結果可能是受到DNA污染的影響[10]。
描述

大角鹿具有所有已知的鹿中最大的鹿角,由鹿角一端至另一端最長可達3.65米(12.0英尺)並重達40(88磅);個體肩高2.1米(6英尺11英寸),體重約為540—600(1,200—1,300磅),最重可超過700(1,500磅)[19][20],最目前已知最重的舊世界鹿科物種之一[8],與現存的阿拉斯加駝鹿同為目前已知第三大型的鹿,第二及最大型的鹿則分別為罕角駝鹿與寬角駝鹿[19][20]。相較於駝鹿,大角鹿具有更為粗壯的骨架,中老年駝鹿的骨架約與壯年大角鹿相當,而壯年駝鹿的骨架則約與青年大角鹿相當。由於社會結構的不同,大角鹿擁有比駝鹿更為明顯的兩性異型[21],雄性的體重約為450—700(990—1,540磅),平均為575(1,268磅),而雌性的體型約為雄性的80%,平均體重為460(1,010磅)[2]。

假設套用與歐洲馬鹿相同的進食模式,一頭擁有40(88磅)鹿角的大角鹿在健康狀況較差時鹿角重量會縮減至20至28(44至62磅)[5][20],而擁有35(77磅)鹿角的大角鹿在狀況差時則會縮減至 18至25(40至55磅)[22],接近現存駝鹿的鹿角重量。另外在氣候條件較差時,擁有35(77磅)鹿角的大角鹿其鹿角重量更可能縮減至低於13(29磅),接近現存加拿大馬鹿的鹿角重量[2]。另外棲息的環境也會影響大角鹿鹿角的形狀,例如棲息於密林中的大角鹿,其鹿角的分支就會較為密集向上[22]。在島嶼上,鹿角基部的圓周大小縮減了41%,按照此比例計算,棲息於島嶼上的大角鹿其鹿角重量僅有棲息於大陸上個體的1/14,這樣的大小差異比起現存歐洲馬鹿更為兩極[5]。大角鹿可能會於繁殖期時脫落並長出新的鹿角。鹿角的生長需要很多的鈣和磷酸鹽,這些同時也是個體維持骨頭必需的營養,也因此當鹿角生長時大角鹿會面臨類似骨質疏鬆症的症狀,大角鹿可以透過食用植物以及脫落的鹿角來重新獲取這些流失的營養[20]。
1998年,加拿大生物學家瓦萊里烏斯‧蓋斯特提出大角鹿為適於奔跑的的動物。他發現到大角鹿的外型類似於現存的馴鹿,而身形比例則類似於善於奔跑的旋角羚、劍羚和高鼻羚羊。共同特徵包括相對較短的四肢、前肢長度與後肢長度近似以及粗壯呈圓柱型的軀體。高鼻羚羊、角馬、馴鹿奔跑速度可超過80 km/h(50 mph)並維持最多15分鐘[2]。
由舊石器時代晚期時期的藝術可知,大角鹿具有較淺的體色,背部顏色較深,於臉頰後及前胸有環帶狀的深色毛髮,軀體兩側則各有一條深色的條紋自肩部延伸至腰腿部,且於馬肩隆處具有駝峰。1989年,美國古生物學家戴爾‧蓋瑟瑞認為,大角鹿的駝峰和野牛一樣,能使前肢擁有更大的轉動幅度,於奔跑時提高步伐的跨距。此外駝峰也能用來儲存脂肪,將脂肪儲存於駝峰而非散布於全身能避免大角鹿於奔跑或夏天發情時過熱[2]。
古生態學
繁殖

在愛爾蘭都柏林郡的巴利貝格(Ballybetagh)酸沼中,發現了超過100具大角鹿的化石,均為只有小角的雄鹿,這代表大角鹿的雄鹿與雌鹿於冬天和春天時是不會混群的;這項特徵與現存很多鹿相同,因為雄鹿與雌鹿有不同的營養需求,並會以不同種類的植物為食。雄雌個體的隔離代表大角鹿可能行一雄多雌的制度,雄鹿會於發情期時互相爭鬥,爭取雌鹿群的交配權。但是於酸沼中發現的大角鹿遺骸均為營養不良的亞成年與老年個體,這代表這群鹿可能死於凍死。目前已知大部分的大角鹿化石可能都來自凍死的個體,而凍死也常常是許多現存鹿的死因。雄鹿多半擁有較高的死亡率,因為牠們在秋天發情期時的進食量較少[23]。在接近發情期時,雄鹿的體重可以由575(1,268磅)提升至690(1,520磅),為了維護龐大身軀所需的能量,加上發情期時的進食量較少,讓雄鹿難以儲存額外足夠的脂肪過冬[2]。
大角鹿的鹿角多半認為是用於繁殖期雄性之間爭鬥時使用[24]、向雌性展示[5]、或是威嚇其他雄性[23]。
現存鹿的妊娠期隨著體型增長而增長,一頭460(1,010磅)的雌性大角鹿其妊娠期約為274天。如果與現存鹿的生存模式近似,大角鹿的鹿角會約於三月上旬時脫落,六月中旬達到最高生長速度,並於七月中旬完成鹿角的生長,七月下旬時鹿茸脫落,發情期的高峰發生於八月的第二週。蓋斯特提出大角鹿是擅於奔跑的動物,也因此他認為雌鹿有能力提供高營養的鹿乳讓幼鹿有足夠的能量與耐力可以跟上鹿群[2]。
棲息地

既非愛爾蘭的特有種也不是駝鹿,大角鹿暱稱為愛爾蘭駝鹿的主要原因是因為目前已知最完整的大角鹿化石均出土自愛爾蘭的湖底沉積或是酸性泥炭沼澤中。大角鹿具有十分廣泛的棲息範圍,最西端可至大西洋,最東則可達貝加爾湖;不過牠們並沒有棲息於北方開闊的猛瑪草原,而主要棲息於北方的林地草原交接帶,主要樹木為分散分布的雲杉與松樹,其他較低矮的植被則包括草、莎草、麻黃和莧[4]。大角鹿的牙齒磨損狀態顯示牠們以多樣的植物為食,會去取食樹葉與嫩枝,或是牧食地面的植被,由牙齒上的花粉殘骸可知道於43,000年前的北海地區主要優勢植物物種為蒿和菊,其他植被則包括車前草、半日花、白花丹與柳樹[25]。針對更新世晚期大角鹿化石所做的同位素分析顯示牠們主要以草和其他非禾本草本植物為食,並於食物較為短缺時透過取食補足營養[26]。透過分析英國更新世中晚期大角鹿的牙齒磨損形式顯示,當地的大角鹿主要也是以牧食為主,但同樣也會取食樹葉[27]。
如果大角鹿營養上的需求與歐洲馬鹿類似,則一頭675(1,488磅)的雄性大角鹿一天需要攝取39.7(88磅)的新鮮植物。若一年中同樣有120天的鹿角成長週期,則每天會需要1,372 g(48.4 oz)的蛋白質,這些蛋白質必須於鹿角開始生長前一個月開始攝食高蛋白或高礦物質含量的植物,直到鹿角生長完成。這樣的植物並不常見,大角鹿可能因此必須前往湖沼並取食其中的水生植物。在鹿角生長完成後,大角鹿可以於由柳樹和樺木圍繞的莎草地獲取足夠的營養[2]。
滅絕


有關大角鹿滅絕的成因討論都主要集中在其鹿角,而非其體型,這可能是由於鹿角的外觀多於其實際用途。有些學者指被人類獵殺是其中一個因素,因大角鹿的鹿角限制了牠們在森林的活動範圍。[5]不過有反駁指大角鹿作為大陸上的物種,應該與人類一同演化,並且適應了人類的存在。
當氣候在最後的冰河時期末改變時,棲息地的植物亦估計改變成為足以提供足夠的養料。在西伯利亞西部發現最近的大角鹿標本(估年約屬於7700年前,已在冰河時期以後),顯示沒有任何養份壓力的徵象。這明顯大角鹿棲息的大陸性氣候地方沒有出現或未曾出現所謂的植物改變。[3]
總的來說,大角鹿在個別地方上消失的成因有多個理論,但就最終大角鹿在歐亞大陸上的消失則沒有定論。有指這是人類存在及棲息地減少聯合造成。[3]
參考
- Lister, A.M. . Quaternary Newsletter. 1987, 52: 14–16.
- Geist, Valerius. . . Stackpole Books. 1998. ISBN 0-8117-0496-3.
- Stuart, A.J.; Kosintsev, P.A.; Higham, T.F.G.; Lister, A.M. (PDF). Nature. 2004, 431 (7009): 684–689 [2007-12-26]. Bibcode:2004Natur.431..684S. PMID 15470427. doi:10.1038/nature02890. (原始内容 (PDF)存档于2006-09-14). Supplementary information (页面存档备份,存于). Erratum in Stuart, A. J. . Nature. 2005, 434 (7031): 413. Bibcode:2005Natur.434..413S. doi:10.1038/nature03413.
- Lister, Adrian M.; Stuart, Anthony J. . Quaternary International. January 2019, 500: 185–203. Bibcode:2019QuInt.500..185L. doi:10.1016/j.quaint.2019.03.025 (英语).
- Gould, Stephen J. . Evolution. 1974, 28 (2): 191–220. doi:10.2307/2407322.
- Hughes, Sandrine; Hayden, Thomas J.; Douady, Christophe J.; Tougard, Christelle; Germonpré, Mietje; Stuart, Anthony; Lbova, Lyudmila; Carden, Ruth F.; Hänni, Catherine; Say, Ludovic. (PDF). Molecular Phylogenetics and Evolution. 2006, 40 (1): 285–291 [2007-12-26]. doi:10.1016/j.ympev.2006.02.004. (原始内容 (PDF)存档于2007-10-01).
- Kuehn, Ralph; Ludt, Christian J.; Schroeder, Wolfgang; Rottmann, Oswald. . Zoological Science. 2005, 22 (9): 1031–1044. PMID 16219984. doi:10.2108/zsj.22.1031.
- Lister, Adrian M.; Edwards, Ceiridwen J.; Nock, D. A. W.; Bunce, Michael; van Pijlen, Iris A.; Bradley, Daniel G.; Thomas, Mark G.; Barnes, Ian. . Nature. 2005, 438 (7069): 850–853. Bibcode:2005Natur.438..850L. PMID 16148942. doi:10.1038/nature04134.
- Immel, Alexander; Drucker, Dorothée G.; Bonazzi, Marion; Jahnke, Tina K.; Münzel, Susanne C.; Schuenemann, Verena J.; Herbig, Alexander; Kind, Claus-Joachim; Krause, Johannes. . Scientific Reports. 2015, 5 (10853): 10853. Bibcode:2015NatSR...510853I. PMC 4459102
 . PMID 26052672. doi:10.1038/srep10853.
. PMID 26052672. doi:10.1038/srep10853. - Mennecart, Bastien; DeMiguel, Daniel; Bibi, Faysal; Rössner, Gertrud E.; Métais, Grégoire; Neenan, James M.; Wang, Shiqi; Schulz, Georg; Müller, Bert; Costeur, Loïc. . Scientific Reports. 13 October 2017, 7 (1): 13176. Bibcode:2017NatSR...713176M. ISSN 2045-2322. PMC 5640792 . PMID 29030580. doi:10.1038/s41598-017-12848-9 (英语).
- Molyneux, T. . Philosophical Transactions of the Royal Society. 1695, 19 (227): 489–512. Bibcode:1695RSPT...19..489M. doi:10.1098/rstl.1695.0083.
- Zimmer, Carl. "The Allure of Big Antlers: The Loom." The Loom. Discover, 3 Sept. 2008. Web. 23 Oct. 2014.
- van der Made, J.; Tong, H.W. . Quaternary International. March 2008, 179 (1): 135–162. Bibcode:2008QuInt.179..135V. doi:10.1016/j.quaint.2007.08.017 (英语).
- Van der Made, Jan. 2019. The dwarfed "giant deer" Megaloceros matritensis n.sp. from the Middle Pleistocene of Madrid - A descendant of M. savini and contemporary to M. giganteus. Quaternary International in press. . Accessed 2019-02-04.
- Lister, Adrian M. . Zoological Journal of the Linnean Society. September 1994, 112 (1–2): 65–100. doi:10.1111/j.1096-3642.1994.tb00312.x (英语).
- Jaubert, Jacques. . OCLC 803593335.
- Johnston, Penny; Kelly, Bernice; Tierney, John. (PDF). Seanda: The NRA Archaeology Magazine: 58–59. [2020-06-02]. (原始内容 (PDF)存档于2013-10-29).
- Kuehn, R.; Ludt, C. J.; Shroeder, W.; Rottmann, O. . Zoological Science. 2005, 22 (9): 1031–1044. PMID 16219984. doi:10.2108/zsj.22.1031.
- R. D. E. Mc Phee, Extinctions in Near Time: Causes, Contexts, and Consequences p.262
- Moen, Ron A.; Pastor, John; Cohen, Yosef. (PDF). Evolutionary Ecology Research. 1999: 235–249 [2020-05-22]. (原始内容存档 (PDF)于2016-03-04).
- Breda, M. . Geobios. 2005, 38 (2): 151–170. doi:10.1016/j.geobios.2003.09.008.
- O'Driscoll Worman, Cedric; Kimbrell, Tristan. . Oikos. 2008, 117 (9): 1397–1405. doi:10.1111/j.0030-1299.2008.16608.x.
- Barnosky, Anthony D. . Science. New. 19 April 1985, 228 (4697): 340–344. Bibcode:1985Sci...228..340B. PMID 17790237. doi:10.1126/science.228.4697.340.
- Kitchener, A. . Modern Geology. 1987, 11: 1–28.
- van Geel, B.; Sevink, J.; Mol, D.; Langeveld, B. W.; van der Ham, R. W. J. M.; van der Kraan, C. J. M.; van der Plicht, J.; Haile, J. S.; Rey-Iglesia, A.; Lorenzen, E. D. . Journal of Quaternary Science. November 2018, 33 (8): 924–933. doi:10.1002/jqs.3069 (英语).
- Chritz, Kendra L.; Dyke, Gareth J.; Zazzo, Antoine; Lister, Adrian M.; Monaghan, Nigel T.; Sigwart, Julia D. . Palaeogeography, Palaeoclimatology, Palaeoecology. November 2009, 282 (1–4): 133–144. Bibcode:2009PPP...282..133C. doi:10.1016/j.palaeo.2009.08.018 (英语).
- Rivals, Florent; Lister, Adrian M. . Quaternary Science Reviews. August 2016, 146: 126. Bibcode:2016QSRv..146..116R. doi:10.1016/j.quascirev.2016.06.007 (英语).
- Stuart, A. J.; Lister, A. M. . Quaternary Science Reviews. 2011, 30 (17–18): 2337. doi:10.1016/j.quascirev.2010.04.023.
- Diedrich, C. G. . Historical Biology. 2011, 24 (2): 161–185. doi:10.1080/08912963.2011.591491.