多智龍屬
多智龙(属名:Tarchia,意为“聪明的”)是生存于晚白垩世蒙古的一属植食性甲龙科恐龙。
| 多智龙属 化石时期:晚白垩世, | |
|---|---|
 | |
| 特氏多智龙正模标本PIN 3142/250的模型 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 科: | †甲龙科 Ankylosauridae |
| 属: | †多智龙属 Tarchia Maryanska, 1977 |
| 模式種 | |
| †凯氏多智龙 Tarchia kielanae Maryanska, 1977 | |
| 其它物种 | |
| |
| 異名 | |
| |
发现与命名
.JPG.webp)
1970年,一支波兰-蒙古考察队在胡山(Khulsan)附近发现一个甲龙类颅骨。1977年,特蕾莎·马瑞沿斯卡(Teresa Maryańska)命名并描述模式种凯氏多智龙(Tarchia kielanae)。属名取自蒙古语 (tarkhi,脑袋)和拉丁语後綴 -ia,指推测其脑部尺寸大于近亲美甲龙。种名纪念考察队的领导者索菲娅·凯兰-伽沃罗斯卡(Zofia Kielan-Jaworowska)教授。正模标本ZPal MgD-I/111发现于蒙古耐梅盖特盆地晚白垩世(可能是坎帕阶至马斯特里赫特阶)的上西戈约特组(原下耐梅盖特层),由一个颅顶、脑壳及颅骨后段材料组成。马瑞沿斯卡将三个额外标本归入该属:ZPAL MgDI/43(一个大型颅后骨骼,由三节单个尾椎、尾槌“柄”的十二节尾椎及一块鳞甲)、ZPAL MgDI/49(一个右肱骨)和PIN 3142/251(一个带有颅骨的骨骼,目前尚未描述)。[1]
多智龙是地质学上已知最年轻的亚洲甲龙科。1977年,塔季扬娜·图曼诺瓦(Tatyana Tumanova)命名第二个种:巨型多智龙(Tarchia gigantea)。这是对巨型倍甲龙(Dyoplosaurus giganteus Maleev 1956)的重新命名,而后者是根据标本PIN 551/29所命名。[2]1987年,图曼诺瓦断定多智龙的两个种完全相同,导致巨型倍甲龙成为凯氏种的首异名。[3]该结论被广泛接受,巨型种也开始作为新组合替代凯氏种成为常用种名。然而维多利亚·阿布尔(Victoria Megan Arbour)的近期研究显示,巨型倍甲龙无法与蒙古晚坎帕阶至马斯特里赫特阶的其它甲龙类进行区分,因此是个疑名;该研究亦恢复凯氏种的名称。[4]
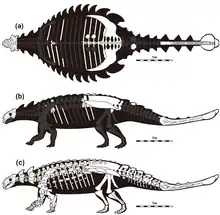
标本ZPAL MgD I/113是一块带有尾部及尾槌的臀部化石,曾先后归入巨型倍甲龙及巨型多智龙,但是阿布尔发现其与巨型倍甲龙正模标本存在差异。[5]这项由阿布尔进行的研究还指出图曼诺瓦于1977年归入智龙的PIN 3142/250标本可能属于美甲龙。这将从根本上改变多智龙的常见外观,因为它是迄今为止保存最完好的多智龙化石,大多数复原图、博物馆骨架及科学研究皆基于该标本。阿布尔发现,多智龙正模标本与牛头怪甲龙(Minotaurasaurus Miles & Miles 2009)存在共同特征,得出结论称后者是前者的次异名。[6]
后来彭卡斯基和图曼诺瓦2016年的一项研究表明,由于PIN 3142/250存在明显解剖学差异,因此不能归入美甲龙,而是代表多智龙的一个新种:特氏多智龙(T. teresae)。 该研究还承认牛头怪甲龙是独立属。[7]2021年,金·帕克(Jin-Young Park)和团队命名了多智龙新种图氏多智龙(T. tumanovae),所知于正模标本MPC-D 100/1353,由一个带有连接颅骨的部分骨骼组成。其发现于赫明察夫(Hermiin Tsav)地区的耐梅盖特组,使之成为特氏种的同时期物种。[8]
描述

多智龙的体型估计主要基于巨型倍甲龙,其正模标本是已知的最大的甲龙类个体之一。这将使多智龙成为已知最长的亚洲甲龙类,身长估计为8(26英尺)。[9]令人困惑的是,经常提及的颅骨尺寸即40(16英寸)长、45(18英寸)宽则基于标本PIN 3142/250――一个小得多的个体。凯氏多智龙与牛头怪甲龙的正模标本也显示出中等体型。2016年,格雷戈里·保罗(Gregory Scott Paul)估计多智龙长5.5(18英尺)、重2.5公噸(390英石),表明牛头怪甲龙是该物种的幼体。[10]
作为甲龙类,多智龙强壮的短腿上应该承载一个宽阔低矮的身体。全身受到皮肤骨化物的保护,称作皮内成骨。它可能拥有骨质尾槌以抵御捕食者。
以前,多智龙可根据较大的基颅骨、未融合的枕旁突-方骨接触面与PIN 3142/250展现的一个事实――即前颌骨的喙宽于上颌齿列最大间距与美甲龙进行区分。2014 年,阿布尔报告两个除仅所知于牛头怪甲龙正模标本以外的显著特征:俯视图中可以看见后脑勺;一个深沟沿鳞状角前侧及外侧延伸,并在前方绕后眶上骨上的附属皮内成骨形成一个深沟。[4]

2016年对多智龙的重新描述指出其与美甲龙的区别在于:具有眶后窝(将鳞状角与眶上骨隔开)和附属皮内成骨;枕骨在背视图中可见;大而深的脑壳;大孔高度大于宽度;颈部皮内成骨外侧高于内侧。此外,其与美甲龙和牛头怪甲龙的区别还在于没有postocular caputegulae(眼眶后面的多边形小骨板),并且尾视图中有成比例增高的枕骨。[7]研究还发现PIN 3142/250(即特氏多智龙)可根据附属皮内成骨未与颅顶融合、方骨未与枕旁突融合、颅顶纹路明显和第四至第十二脑神经孔分叉与凯氏多智龙区分。[7]
较早文献中给出关于多智龙的许多信息皆参考PIN 3142/250(2016年命名为特氏多智龙之前曾简单归入美甲龙)。2001年,有人指出有证据表明多智龙拥有牙齿咬合磨损面;[11]这可能不是在说正模标本,因为正模标本中没有保存牙齿。
种系发生学
维卡尤斯等人2004年指出,多智龙处在晚白垩世甲龙科两个不同演化支的基干位置:一个包括北美分类单元(甲龙、包头龙),另一个包括亚洲分类单元(绘龙未定种、美甲龙、天镇龙、篮尾龙)。[12]然而,此结论又是基于PIN 3142/250标本,其特征通常在各种支序分类学分析中定义为多智龙的操作分类单元。值得注意的是,在这些分析中,多智龙和美甲龙经常处于非常不同的位置。
以下分支图基于阿布尔和柯里2015年的甲龙科系统发育分析:[13]
| 甲龙亚科 Ankylosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2016年多智龙重新描述中进行的有限系统发育分析重点关注多智龙、美甲龙和牛头怪甲龙之间的相互关系,复制如下。[7]
| ||||||||||||||||||||||||||||
参见
- 甲龙类研究历史
- 2017年主龙类古生物学研究
参考资料
- Maryańska, T. . Palaeontologia Polonica. 1977, 37: 85–151.
- Tumanova, T. A. . Paleontological Journal. 1977, 11: 480–486.
- T. A. Tumanova, 1987, Pantsirnyye dinozavry Mongolii, Trudy Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya 32, 80 pp
- Arbour, Victoria Megan, 2014. Systematics, evolution, and biogeography of the ankylosaurid dinosaurs. Ph.D thesis, University of Alberta. https://era.library.ualberta.ca/public/.../Arbour_Victoria_Spring2014.pdf%5B%5D
- Victoria M. Arbour; Nicolai L. Lech-Hernes; Tom E. Guldberg; Jørn H. Hurum; Philip J. Currie. . Acta Palaeontologica Polonica. 2013, 58: 55–64.
- Victoria M. Arbour; Philip J. Currie; Demchig Badamgarav. . Zoological Journal of the Linnean Society. 2014, 172 (3): 631–652.
- Paul Penkalski; Tatiana Tumanova. . Cretaceous Research. 2016, 70: 117–127. doi:10.1016/j.cretres.2016.10.004.
- Park JY, Lee YN, Kobayashi Y, Jacobs LL, Barsbold R, Lee HJ, Kim N, Song KY, Polcyn MJ. . Scientific Reports. 2021, 11 (1): Article number 22928. PMC 8616956
 . PMID 34824329. doi:10.1038/s41598-021-02273-4 .
. PMID 34824329. doi:10.1038/s41598-021-02273-4 . - Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix. (页面存档备份,存于)
- Paul, G. S. . Princeton University Press. 2016: 261.
- Barrett, P. M. . Carpenter, K. (编). . Bloomington: Indiana University Press. 2001: 25–52.
- Vickaryous, Maryańska, and Weishampel. . Weishampel, D. B.; Dodson, P.; Osmólska, H. (编). . University of California Press. 2004.
- Arbour, V. M.; Currie, P. J. . Journal of Systematic Palaeontology. 2015, 14 (5): 1–60. S2CID 214625754. doi:10.1080/14772019.2015.1059985.
- Holtz, Jr., Thomas.
 . New York, New York: Random House, Inc. 2007: 241. ISBN 9780375824197.
. New York, New York: Random House, Inc. 2007: 241. ISBN 9780375824197. - Gallagher, W. B.; Tumanova, T. A.; Dodson, P.; Axel, L. . Journal of Vertebrate Paleontology. 1998, 18: 44A–45A.